
Why Darwinism Is False
Jerry A. Coyne is a professor in the Department of Ecology and Evolution at The University of Chicago. In Why Evolution is True, he summarizes Darwinism — the modern theory of evolution — as follows: “Life on earth evolved gradually beginning with one primitive species — perhaps a self-replicating molecule — that lived more than 3.5 billion years ago; it then branched out over time, throwing off many new and diverse species; and the mechanism for most (but not all) of evolutionary change is natural selection.”1
Coyne further explains that evolution “simply means that a species undergoes genetic change over time. That is, over many generations a species can evolve into something quite different, and those differences are based on changes in the DNA, which originate as mutations. The species of animals and plants living today weren’t around in the past, but are descended from those that lived earlier.”2
According to Coyne, however, “if evolution meant only gradual genetic change within a species, we’d have only one species today — a single highly evolved descendant of the first species. Yet we have many… How does this diversity arise from one ancestral form?” It arises because of “splitting, or, more accurately, speciation,” which “simply means the evolution of different groups that can’t interbreed.”3
If Darwinian theory were true, “we should be able to find some cases of speciation in the fossil record, with one line of descent dividing into two or more. And we should be able to find new species forming in the wild.” Furthermore, “we should be able to find examples of species that link together major groups suspected to have common ancestry, like birds with reptiles and fish with amphibians.” Finally, there are facts that “make sense only in light of the theory of evolution” but do not make sense in the light of creation or design. These include “patterns of species distribution on the earth’s surface, peculiarities of how organisms develop from embryos, and the existence of vestigial features that are of no apparent use.” Coyne concludes his introduction with the bold statement that “all the evidence — both old and new — leads ineluctably to the conclusion that evolution is true.”4
Of course, “evolution” is undeniably true if it means simply that existing species can change in minor ways over time, or that many species living today did not exist in the past. But Darwin’s claim that all species are modified descendants of a common ancestor, and Coyne’s claim that DNA mutations and natural selection have produced those modifications, are not so undeniably true. Coyne devotes the remainder of his book to providing evidence for them.
Fossils
Coyne turns first to the fossil record. “We should be able,” he writes, “to find some evidence for evolutionary change in the fossil record. The deepest (and oldest) layers of rock would contain the fossils of more primitive species, and some fossils should become more complex as the layers of rock become younger, with organisms resembling present-day species found in the most recent layers. And we should be able to see some species changing over time, forming lineages showing ‘descent with modification’ (adaptation).” In particular, “later species should have traits that make them look like the descendants of earlier ones.”5
In The Origin of Species, Charles Darwin acknowledged that the fossil record presented difficulties for his theory. “By the theory of natural selection,” he wrote, “all living species have been connected with the parent-species of each genus, by differences not greater than we see between the natural and domestic varieties of the same species at the present day.” Thus in the past “the number of intermediate and transitional links, between all living and extinct species, must have been inconceivably great.” But Darwin knew that the major animal groups — which modern biologists call “phyla” — appeared fully formed in what were at the time the earliest known fossil-bearing rocks, deposited during a geological period known as the Cambrian. He considered this a “serious” difficulty for his theory, since “if the theory be true, it is indisputable that before the lowest Cambrian stratum was deposited long periods elapsed… and that during these vast periods the world swarmed with living creatures.” And “to the question why we do not find rich fossiliferous deposits belonging to these assumed earliest periods prior to the Cambrian system, I can give no satisfactory answer.” So “the case at present must remain inexplicable; and may be truly urged as a valid argument against the views here entertained.”6
Darwin defended his theory by citing the imperfection of the geological record. In particular, he argued that Precambrian fossils had been destroyed by heat, pressure, and erosion. Some of Darwin’s modern followers have likewise argued that Precambrian fossils existed but were later destroyed, or that Precambrian organisms were too small or too soft to have fossilized in the first place. Since 1859, however, paleontologists have discovered many Precambrian fossils, many of them microscopic or soft-bodied. As American paleobiologist William Schopf wrote in 1994, “The long-held notion that Precambrian organisms must have been too small or too delicate to have been preserved in geological materials… [is] now recognized as incorrect.” If anything, the abrupt appearance of the major animal phyla about 540 million years ago — which modern biologists call “the Cambrian explosion” or “biology’s Big Bang” — is better documented now than in Darwin’s time. According to Berkeley paleontologist James Valentine and his colleagues, the “explosion is real, it is too big to be masked by flaws in the fossil record.” Indeed, as more fossils are discovered it becomes clear that the Cambrian explosion was “even more abrupt and extensive than previously envisioned.”7
What does Coyne’s book have to say about this?
“Around 600 million years ago,” Coyne writes, “a whole gamut of relatively simple but multicelled organisms arise, including worms, jellyfish, and sponges. These groups diversify over the next several million years, with terrestrial plants and tetrapods (four-legged animals, the earliest of which were lobe-finned fish) appearing about 400 million years ago.”8
In other words, Coyne’s account of evolutionary history jumps from 600 to 400 million years ago without mentioning the 540 million year-old Cambrian explosion. In this respect, Coyne’s book reads like a modern biology textbook that has been written to indoctrinate students in Darwinian evolution rather than provide them with the facts.
Coyne goes on to discuss several “transitional” forms. “One of our best examples of an evolutionary transition,” he writes, is the fossil record of whales, “since we have a chronologically ordered series of fossils, perhaps a lineage of ancestors and descendants, showing their movement from land to water.”9
“The sequence begins,” Coyne writes, “with the recently discovered fossil of a close relative of whales, a raccoon-sized animal called Indohyus. Living 48 million years ago, Indohyus was… probably very close to what the whale ancestor looked like.” In the next paragraph, Coyne writes, “Indohyus was not the ancestor of whales, but was almost certainly its cousin. But if we go back 4 million more years, to 52 million years ago, we see what might well be that ancestor. It is a fossil skull from a wolf-sized creature called Pakicetus, which is bit more whalelike than Indohyus.” On the page separating these two paragraphs is a figure captioned “Transitional forms in the evolution of modern whales,” which shows Indohyus as the first in the series and Pakicetus as the second.10
But Pakicetus — as Coyne just told us — is 4 million years older than Indohyus. To a Darwinist, this doesn’t matter: Pakicetus is “more whalelike” than Indohyus, so it must fall between Indohyus and modern whales, regardless of the fossil evidence.
(Coyne performs the same trick with fossils that are supposedly ancestral to modern birds. The textbook icon Archaeopteryx, with feathered wings like a modern bird but teeth and a tail like a reptile, is dated at 145 million years. But what Coyne calls the “nonflying feathered dinosaur fossils” — which should have come before Archaeopteryx — are tens of millions of years younger. Like Darwinists Kevin Padian and Luis Chiappe eleven years earlier, Coyne simply rearranges the evidence to fit Darwinian theory.)11
So much for Coyne’s prediction that “later species should have traits that make them look like the descendants of earlier ones.” And so much for his argument that “if evolution were not true, fossils would not occur in an order that makes evolutionary sense.” Ignoring the facts he himself has just presented, Coyne brazenly concludes: “When we find transitional forms, they occur in the fossil record precisely where they should.” If Coyne’s book were turned into a movie, this scene might feature Chico Marx saying, “Who are you going to believe, me or your own eyes?”12
There is another problem with the whale series (and every other series of fossils) that Coyne fails to address: No species in the series could possibly be the ancestor of any other, because all of them possess characteristics they would first have to lose before evolving into a subsequent form. This is why the scientific literature typically shows each species branching off a supposed lineage.
In the figure below, all the lines are hypothetical. The diagram on the left is a representation of evolutionary theory: Species A is ancestral to B, which is ancestral to C, which is ancestral to D, which is ancestral to E. But the diagram on the right is a better representation of the evidence: Species A, B, C and D are not in the actual lineage leading to E, which remains unknown.
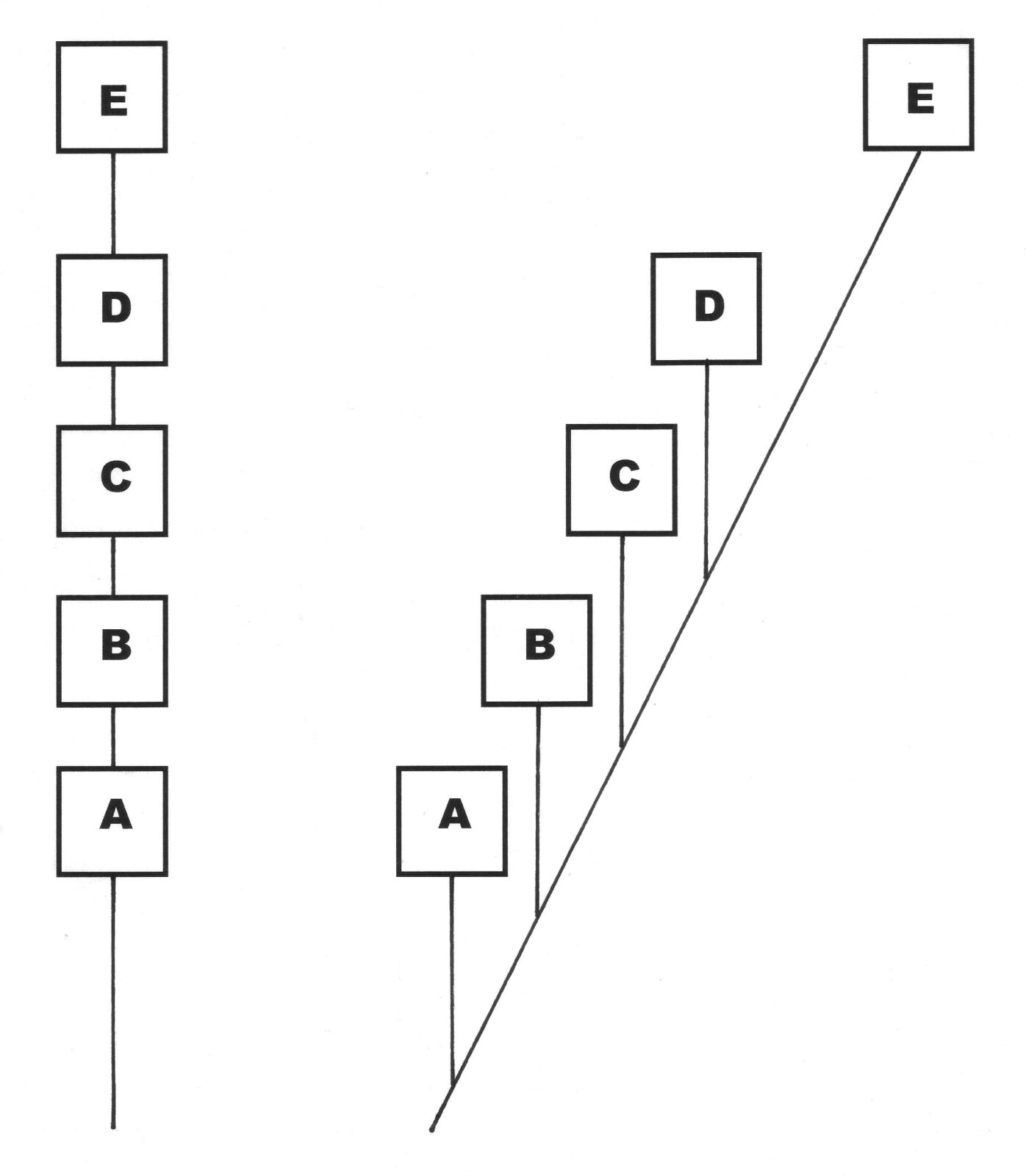
It turns out that no series of fossils can provide evidence for Darwinian descent with modification. Even in the case of living species, buried remains cannot generally be used to establish ancestor-descendant relationships. Imagine finding two human skeletons in the same grave, one about thirty years older than the other. Was the older individual the parent of the younger? Without written genealogical records and identifying marks (or in some cases DNA), it is impossible to answer the question. And in this case we would be dealing with two skeletons from the same species that are only a generation apart and from the same location. With fossils from different species that are now extinct, and widely separated in time and space, there is no way to establish that one is the ancestor of another—no matter how many transitional fossils we find.
In 1978, Gareth Nelson of the American Museum of Natural History wrote: “The idea that one can go to the fossil record and expect to empirically recover an ancestor-descendant sequence, be it of species, genera, families, or whatever, has been, and continues to be, a pernicious illusion.”13 Nature science writer Henry Gee wrote in 1999 that “no fossil is buried with its birth certificate.” When we call new fossil discoveries “missing links,” it is “as if the chain of ancestry and descent were a real object for our contemplation, and not what it really is: a completely human invention created after the fact, shaped to accord with human prejudices.” Gee concluded: “To take a line of fossils and claim that they represent a lineage is not a scientific hypothesis that can be tested, but an assertion that carries the same validity as a bedtime story — amusing, perhaps even instructive, but not scientific.”14
Embryos
So evolutionary theory needs better evidence than the fossil record can provide. Coyne correctly notes: “When he wrote The Origin, Darwin considered embryology his strongest evidence for evolution.” Darwin had written that the evidence seemed to show that “the embryos of the most distinct species belonging to the same class are closely similar, but become, when fully developed, widely dissimilar,” a pattern that “reveals community of descent.” Indeed, Darwin thought that early embryos “show us, more or less completely, the condition of the progenitor of the whole group in its adult state.”15
But Darwin was not an embryologist. In The Origin of Species he supported his contention by citing a passage by German embryologist Karl Ernst von Baer:
“The embryos of mammals, birds, lizards and snakes, and probably chelonia [turtles] are in their earliest states exceedingly like one another. … In my possession are two little embryos in spirit, whose names I have omitted to attach, and at present I am quite unable to say to what class they belong. They may be lizards or small birds, or very young mammals, so complete is the similarity in the mode of formation of the head and trunk in these animals.”16
Coyne claims that this is something von Baer “wrote to Darwin,” but Coyne’s history is as unreliable as his paleontology. The passage Darwin cited was from a work written in German by von Baer in 1828; Thomas Henry Huxley translated it into English and published it in 1853. Darwin didn’t even realize at first that it was from von Baer: In the first two editions of The Origin of Species he incorrectly attributed the passage to Louis Agassiz.17
Ironically, von Baer was a strong critic of Darwin’s theory, rejecting the idea that all vertebrates share a common ancestor. According to historian of science Timothy Lenoir, von Baer feared that Darwin and his followers had “already accepted the Darwinian evolutionary hypothesis as true before they set to the task of observing embryos.” The myth that von Baer’s work supported Darwin’s theory was due primarily to another German biologist, Ernst Haeckel.”18 Haeckel maintained not only that all vertebrate embryos evolved from a common ancestor, but also that in their development (“ontogeny”) they replay (“recapitulate”) their evolutionary history (“phylogeny”). He called this The Biogenetic Law: Ontogeny recapitulates phylogeny.
In Why Evolution Is True, Coyne writes that “the ‘recapitulation’ of an evolutionary sequence is seen in the developmental sequence” of various organs. “Each vertebrate undergoes development in a series of stages, and the sequence of those stages happens to follow the evolutionary sequence of its ancestors.” The probable reason for this is that “as one species evolves into another, the descendant inherits the developmental program of its ancestor.” So the descendant tacks changes “onto what is already a robust and basic developmental plan. It is best for things that evolved later to be programmed to develop later in the embryo. This ‘adding new stuff onto old’ principle also explains why the sequence of developmental stages mirrors the evolutionary sequence of organisms. As one group evolves from another, it often adds its developmental program on top of the old one.” Thus “all vertebrates begin development looking like embryonic fish because we all descended from a fishlike ancestor.”19
Nevertheless, Coyne writes, Haeckel’s Biogenetic Law “wasn’t strictly true,” because “embryonic stages don’t look like the adult forms of their ancestors,” as Haeckel (and Darwin) believed, “but like the embryonic forms of their ancestors.” But this reformulation of The Biogenetic Law doesn’t solve the problem. First, fossil embryos are extremely rare,20 so the reformulated law has to rely on embryos of modern organisms that are assumed to resemble ancestral forms. The result is a circular argument: According to Darwin’s theory, fish are our ancestors; human embryos (allegedly) look like fish embryos; therefore, human embryos look like the embryos of our ancestors. Theory first, observation later — just as von Baer had objected.
Second, the idea that later evolutionary stages can simply be tacked onto development is biologically unrealistic. A human is not just a fish embryo with some added features. As British embryologist Walter Garstang pointed out in 1922, “a house is not a cottage with an extra story on the top. A house represents a higher grade in the evolution of a residence, but the whole building is altered — foundations, timbers, and roof — even if the bricks are the same.”21
Third, and most important, vertebrate embryos are not most similar in their earliest stages. In the 1860s, Haeckel made some drawings to show that vertebrate embryos look almost identical in their first stage — but his drawings were faked. Not only had he distorted the embryos by making them appear more similar than they really are, but he had also omitted earlier stages in which the embryos are strikingly different from each other. A human embryo in its earliest stages looks nothing like a fish embryo.
Only after vertebrate embryos have progressed halfway through their development do they reach the stage that Darwin and Haeckel treated as the first. Developmental biologists call this different-similar-different pattern the “developmental hourglass.” Vertebrate embryos do not resemble each other in their earliest stages, but they converge somewhat in appearance midway through development before diverging again. If ontogeny were a recapitulation of phylogeny, such a pattern would be more consistent with separate origins than with common ancestry. Modern Darwinists attempt to salvage their theory by assuming that the common ancestry of vertebrates is obscured because early development can evolve easily, but there is no justification for this assumption other than the theory itself.22
Although Haeckel’s drawings were exposed as fakes by his own contemporaries, biology textbooks used them throughout the twentieth century to convince students that humans share a common ancestor with fish. Then, in 1997, a scientific journal published an article comparing photos of vertebrate embryos to Haeckel’s drawings, which the lead author described as “one of the most famous fakes in biology.” In 2000, Harvard evolutionary biologist Stephen Jay Gould called Haeckel’s drawings “fraudulent” and wrote that biologists should “be both astonished and ashamed by the century of mindless recycling that has led to the persistence of these drawings in a large number, if not a majority, of modern textbooks.”23
But Coyne is not ashamed. He defends Haeckel’s drawings “Haeckel was accused, largely unjustly,” Coyne writes, “of fudging some drawings of early embryos to make them look more similar than they really are. Yet we shouldn’t throw out the baby with the bath water.”24 The “baby” is Darwin’s theory, which Coyne stubbornly defends regardless of the evidence.
Vestiges and Bad Design
Darwin argued in The Origin of Species that the widespread occurrence of vestigial organs—organs that may have once had a function but are now useless—is evidence against creation. “On the view of each organism with all its separate parts having been specially created, how utterly inexplicable is it that organs bearing the plain stamp of inutility… should so frequently occur.” But such organs, he argued, are readily explained by his theory: “On the view of descent with modification, we may conclude that the existence of organs in a rudimentary, imperfect, and useless condition, or quite aborted, far from presenting a strange difficulty, as they assuredly do on the old doctrine of creation, might even have been anticipated in accordance with the views here explained.”25
In The Descent of Man, Darwin cited the human appendix as an example of a vestigial organ. But Darwin was mistaken: The appendix is now known to be an important source of antibody-producing blood cells and thus an integral part of the human immune system. It may also serve as a compartment for beneficial bacteria that are needed for normal digestion. So the appendix is not useless at all.26
In 1981, Canadian biologist Steven Scadding argued that although he had no objection to Darwinism, “vestigial organs provide no evidence for evolutionary theory.” The primarily reason is that “it is difficult, if not impossible, to unambiguously identify organs totally lacking in function.” Scadding cited the human appendix as an organ previously thought to be vestigial but now known to have a function. Another Canadian biologist, Bruce Naylor, countered that an organ with some function can still be considered vestigial. Furthermore, Naylor argued, “perfectly designed organisms necessitated the existence of a creator,” but “organisms are often something less than perfectly designed” and thus better explained by evolution. Scadding replied: “The entire argument of Darwin and others regarding vestigial organs hinges on their uselessness and inutility.” Otherwise, the argument from vestigiality is nothing more than an argument from homology, and “Darwin treated these arguments separately recognizing that they were in fact independent.” Scadding also objected that Naylor’s “less than perfectly designed” argument was “based on a theological assumption about the nature of God, i.e. that he would not create useless structures. Whatever the validity of this theological claim, it certainly cannot be defended as a scientific statement, and thus should be given no place in a scientific discussion of evolution.”27
In Why Evolution Is True, Coyne (like Darwin) cites the human appendix as an example of a vestigial organ. Unlike Darwin, however, Coyne concedes that “it may be of some small use. The appendix contains patches of tissue that may function as part of the immune system. It has also been suggested that it provides a refuge for useful gut bacteria. But these minor benefits are surely outweighed by the severe problems that come with the human appendix.” In any case, Coyne argues, “the appendix is still vestigial, for it no longer performs the function for which it evolved.”28
As Scadding had pointed out nearly thirty years ago, however, Darwin’s argument rested on lack of function, not change of function. Furthermore, if vestigiality were redefined as Coyne proposes, it would include many features never before thought to be vestigial. For example, if the human arm evolved from the leg of a four-footed mammal (as Darwinists claim), then the human arm is vestigial. And if (as Coyne argues) the wings of flying birds evolved from feathered forelimbs of dinosaurs that used them for other purposes, then the wings of flying birds are vestigial. This is the opposite of what most people mean by “vestigial.”29
Coyne also ignores Scadding’s other criticism, arguing that whether the human appendix is useless or not, it is an example of imperfect or bad design. “What I mean by ‘bad design’,” Coyne writes, “is the notion that if organisms were built from scratch by a designer — one who used the biological building blocks or nerves, muscles, bone, and so on — they would not have such imperfections. Perfect design would truly be the sign of a skilled and intelligent designer. Imperfect design is the mark of evolution; in fact, it’s precisely what we expect from evolution.”30
An even better example of bad design, Coyne argues, is the prevalence of “dead genes.” According to the modern version of Darwinism that Coyne defends, DNA carries a genetic program that encodes proteins that direct embryo development; mutations occasionally alter the genetic program to produce new proteins (or change their locations); and natural selection then sorts those mutations to produce evolution. In the 1970s, however, molecular biologists discovered that most of our DNA does not encode proteins. In 1972 Susumu Ohno called this “junk,” and in 1976 Richard Dawkins wrote: “A large fraction of the DNA is never translated into protein. From the point of view of the individual organism this seems paradoxical. If the ‘purpose’ of DNA is to supervise the building of bodies, it is surprising to find a large quantity of DNA which does no such thing.” From the point of view of Darwinian evolution, however, there is no paradox. “The true ‘purpose’ of DNA is to survive, no more and no less. The simplest way to explain the surplus DNA is to suppose that it is a parasite, or at best a harmless but useless passenger, hitching a ride in the survival machines created by the other DNA.”31
Like Dawkins, Coyne regards much of our DNA as parasitic. He writes in Why Evolution Is True: “When a trait is no longer used, or becomes reduced, the genes that make it don’t instantly disappear from the genome: evolution stops their action by inactivating them, not snipping them out of the DNA. From this we can make a prediction. We expect to find, in the genomes of many species, silenced, or ‘dead,’ genes: genes that once were useful but are no longer intact or expressed. In other words, there should be vestigial genes. In contrast, the idea that all species were created from scratch predicts that no such genes would exist.” Coyne continues: “Thirty years ago we couldn’t test this prediction because we had no way to read the DNA code. Now, however, it’s quite easy to sequence the complete genome of species, and it’s been done for many of them, including humans. This gives us a unique tool to study evolution when we realize that the normal function of a gene is to make a protein—a protein whose sequence of amino acids is determined by the sequence of nucleotide bases that make up the DNA. And once we have the DNA sequence of a given gene, we can usually tell if it is expressed normally — that is, whether it makes a functional protein — or whether it is silenced and makes nothing. We can see, for example, whether mutations have changed the gene so that a usable protein can no longer be made, or whether the ‘control’ regions responsible for turning on a gene have been inactivated. A gene that doesn’t function is called a pseudogene. And the evolutionary prediction that we’ll find pseudogenes has been fulfilled — amply. Virtually every species harbors dead genes, many of them still active in its relatives. This implies that those genes were also active in a common ancestor, and were killed off in some descendants but not in others. Out of about thirty thousand genes, for example, we humans carry more than two thousand pseudogenes. Our genome — and that of other species — are truly well populated graveyards of dead genes.”32
But Coyne is dead wrong.
Evidence pouring in from genome-sequencing projects shows that virtually all of an organism’s DNA is transcribed into RNA, and that even though most of that RNA is not translated into proteins, it performs essential regulatory functions. Every month, science journals publish articles describing more such functions. And this is not late-breaking news: The evidence has been accumulating since 2003 (when scientists finished sequencing the human genome) that “pseudogenes” and other so-called “junk DNA” sequences are not useless after all.33 Why Evolution Is True ignores this enormous body of evidence, which decisively refutes Coyne’s Darwinian prediction that our genome should contain lots of “dead” DNA. It’s no wonder that Coyne falls back again and again on the sort of theological arguments that Scadding wrote “should be given no place in a scientific discussion of evolution.”
Biogeography
Theological arguments are also prominent in The Origin of Species. For example, Darwin argued that the geographic distribution of living things made no sense if species had been separately created, but it did make sense in the context of his theory. Cases such as “the presence of peculiar species of bats on oceanic islands and the absence of all other terrestrial mammals,” Darwin wrote, “are facts utterly inexplicable on the theory of independent acts of creation.” In particular: “Why, it may be asked, has the supposed creative force produced bats and no other mammals on remote islands?” According to Darwin, “on my view this question can easily be answered; for no terrestrial mammal can be transported across a wide space of sea, but bats can fly across.”34
But Darwin knew that migration cannot account for all patterns of geographic distribution. He wrote in The Origin of Species that “the identity of many plants and animals, on mountain-summits, separated from each other by hundreds of miles of lowlands, where Alpine species could not possibly exist, is one of the most striking cases known of the same species living at distant points without the apparent possibility of their having migrated from one point to the other.” Darwin argued that the recent ice age “affords a simple explanation of these facts.” Arctic plants and animals that were “nearly the same” could have flourished everywhere in Europe and North America, but “when the warmth had fully returned, the same species, which had lately lived together on the European and North American lowlands, would again be found in the arctic regions of the Old and New Worlds, and on many isolated mountain-summits far distant from each other.”35
So some cases of geographic distribution may not be due to migration, but to the splitting of a formerly large, widespread population into small, isolated populations — what modern biologists call “vicariance.” Darwin argued that all modern distributions of species could be explained by these two possibilities. Yet there are many cases of geographic distribution that neither migration nor vicariance seem able to explain.
One example is the worldwide distribution of flightless birds, or “ratites.” These include ostriches in Africa, rheas in South America, emus and cassowaries in Australia, and kiwis in New Zealand. Since the birds are flightless, explanations based on migration over vast oceanic distances are implausible. After continental drift was discovered in the twentieth century, it was thought that the various populations might have separated with the landmasses. But ostriches and kiwis are much too recent; the continents had already drifted apart when these species originated. So neither migration nor vicariance explain ratite biogeography.36
Another example is freshwater crabs. Studied intensively by Italian biologist Giuseppe Colosi in the 1920s, these animals complete their life cycles exclusively in freshwater habitats and are incapable of surviving prolonged exposure to salt water. Today, very similar species are found in widely separated lakes and rivers in Central and South America, Africa, Madagascar, southern Europe, India, Asia and Australia. Fossil and molecular evidence indicates that these animals originated long after the continents separated, so their distribution is inconsistent with the vicariance hypothesis. Some biologists speculate that the crabs may have migrated by “transoceanic rafting” in hollow logs, but this seems unlikely given their inability to tolerate salt water. So neither vicariance nor migration provides a convincing explanation for the biogeography of these animals.37
An alternative explanation was suggested in the mid-twentieth century by Léon Croizat, a French biologist raised in Italy. Croizat found that Darwin’s theory did “not seem to agree at all with certain important facts of nature,” especially the facts of biogeography. Indeed, he concluded, “Darwinism is by now only a straitjacket… a thoroughly decrepit skin to hold new wine.” Croizat did not argue for independent acts of creation; instead, he proposed that in many cases a widespread primitive species was split into fragments, then its remnants evolved in parallel, in separate locations, into new species that were remarkably similar. Croizat called this process of parallel evolution “orthogenesis.” Neo-Darwinists such as Ernst Mayr, however, pointed out that there is no mechanism for orthogenesis, which implies — contrary to Darwinism — that evolution is guided in certain directions; so they rejected Croizat’s hypothesis.38
In Why Evolution Is True, Coyne (like Darwin) attributes the biogeography of oceanic islands to migration, and certain other distributions to vicariance. But Coyne (unlike Darwin) acknowledges that these two processes cannot explain everything. For example, the internal anatomy of marsupial mammals is so different from the internal anatomy of placental mammals that the two groups are thought to have split a long time ago. Yet there are marsupial flying squirrels, anteaters and moles in Australia that strikingly resemble placental flying squirrels, anteaters and moles on other continents, and these forms originated long after the continents had separated.
Coyne attributes the similarities to “a well-known process called convergent evolution.” According to Coyne. “It’s really quite simple. Species that live in similar habitats will experience similar selection pressures from their environment, so they may evolve similar adaptations, or converge, coming to look and behave very much alike even though they are unrelated.” Put together common ancestry, natural selection, and the origin of species (“speciation”), “add in the fact that distant areas of the world can have similar habitats, and you get convergent evolution — and a simple explanation of a major geographic pattern.”39
This is not the same as Croizat’s “orthogenesis,” according to which populations of a single species, after becoming separated from each other, evolve in parallel due to some internal directive force. According to Coyne’s “convergent evolution,” organisms that are fundamentally different from each other evolve through natural selection to become superficially similar because they inhabit similar environments. The mechanism for orthogenesis is internal, whereas the mechanism for convergence is external. In both cases, however, mechanism is crucial: Without it, orthogenesis and convergence are simply words describing biogeographical patterns, not explanations of how those patterns originated.
So the same question can be asked of convergence that was asked of orthogenesis: What is the evidence for the proposed mechanism? According to Coyne, the mechanism of convergence involves natural selection and speciation.
Selection and Speciation
Coyne writes that Darwin “had little direct evidence for selection acting in natural populations.” Actually, Darwin had no direct evidence for natural selection; the best he could do in The Origin of Species was “give one or two imaginary illustrations.” It wasn’t until a century later that Bernard Kettlewell provided what he called “Darwin’s missing evidence” for natural selection — a shift in the proportion of light- and dark-colored peppered moths that Kettlewell attributed to camouflage and bird predation.40
Since then, biologists have found lots of direct evidence for natural selection. Coyne describes some of it, including an increase in average beak depth of finches on the Galápagos Islands and a change in flowering time in wild mustard plants in Southern California — both due to drought. Like Darwin, Coyne also compares natural selection to the artificial selection used in plant and animal breeding.
But these examples of selection — natural as well as artificial — involve only minor changes within existing species. Breeders were familiar with such changes before 1859, which is why Darwin did not write a book titled How Existing Species Change Over Time; he wrote a book titled The Origin of Species by Means of Natural Selection. “Darwin called his great work On the Origin of Species,” wrote Harvard evolutionary biologist Ernst Mayr in 1982, “for he was fully conscious of the fact that the change from one species into another was the most fundamental problem of evolution.” Yet, Mayr had written earlier, “Darwin failed to solve the problem indicated by the title of his work.” In 1997, evolutionary biologist Keith Stewart Thomson wrote: “A matter of unfinished business for biologists is the identification of evolution’s smoking gun,” and “the smoking gun of evolution is speciation, not local adaptation and differentiation of populations.” Before Darwin, the consensus was that species can vary only within certain limits; indeed, centuries of artificial selection had seemingly demonstrated such limits experimentally. “Darwin had to show that the limits could be broken,” wrote Thomson, “so do we.”41
In 2004, Coyne and H. Allen Orr published a detailed book titled Speciation, in which they noted that biologists have not been able to agree on a definition of “species” because no single definition fits every case. For example, a definition applicable to living, sexually reproducing organisms might make no sense when applied to fossils or bacteria. In fact, there are more than 25 definitions of “species.” What definition is best? Coyne and Orr argued that, “when deciding on a species concept, one should first identify the nature of one’s ‘species problem,’ and then choose the concept best at solving that problem.” Like most other Darwinists, Coyne and Orr favor Ernst Mayr’s “biological species concept” (BSC), according to which “species are groups of interbreeding natural populations that are reproductively isolated from other such groups.” In Why Evolution Is True, Coyne explains that the biological species concept is “the one that evolutionists prefer when studying speciation, because it gets you to the heart of the evolutionary question. Under the BSC, if you can explain how reproductive barriers evolve, you’ve explained the origin of species.”42
Theoretically, reproductive barriers arise when geographically separated populations diverge genetically. But Coyne describes five “cases of real-time speciation” that involve a different mechanism: chromosome doubling, or “polyploidy.”43 This usually follows hybridization between two existing plant species. Most hybrids are sterile because their mismatched chromosomes can’t separate properly to produce fertile pollen and ovaries; occasionally, however, the chromosomes in a hybrid spontaneously double, producing two perfectly matched sets and making reproduction possible. The result is a fertile plant that is reproductively isolated from the two parents—a new species, according to the BSC.
But speciation by polyploidy (“secondary speciation”) has been observed only in plants. It does not provide evidence for Darwin’s theory that species originate through natural selection, nor for the neo-Darwinian theory of speciation by geographic separation and genetic divergence. Indeed, according to evolutionary biologist Douglas J. Futuyma, polyploidy “does not confer major new morphological characteristics… [and] does not cause the evolution of new genera” or higher levels in the biological hierarchy.44
So secondary speciation does not solve Darwin’s problem. Only primary speciation — the splitting of one species into two by natural selection — would be capable of producing the branching-tree pattern of Darwinian evolution. But no one has ever observed primary speciation. Evolution’s smoking gun has never been found.45
Or has it?
In Why Evolution Is True, Coyne claims that primary speciation was observed in an experiment reported in 1998. Curiously, Coyne did not mention it in the 2004 book he co-authored with Orr, but his 2009 account of it is worth quoting in full:
“We can even see the origin of a new, ecologically diverse bacterial species, all within a single laboratory flask. Paul Rainey and his colleagues at Oxford University placed a strain of the bacteria Pseudomonas fluorescens in a small vessel containing nutrient broth, and simply watched it. (It’s surprising but true that such a vessel actually contains diverse environments. Oxygen concentration, for example, is highest on the top and lowest on the bottom.) Within ten days — no more than a few hundred generations — the ancestral free-floating ‘smooth’ bacterium had evolved into two additional forms occupying different parts of the beaker. One, called ‘wrinkly spreader,’ formed a mat on top of the broth. The other, called ‘fuzzy spreader,’ formed a carpet on the bottom. The smooth ancestral type persisted in the liquid environment in the middle. Each of the two new forms was genetically different from the ancestor, having evolved through mutation and natural selection to reproduce best in their respective environments. Here, then, is not only evolution but speciation occurring in the lab: the ancestral form produced, and coexisted with, two ecologically different descendants, and in bacteria such forms are considered distinct species. Over a very short time, natural selection in Pseudomonas yielded a small-scale ‘adaptive radiation,’ the equivalent of how animals or plants form species when they encounter new environments on an oceanic island.”46
But Coyne omits the fact that when the ecologically different forms were placed back into the same environment, they “suffered a rapid loss of diversity,” according to Rainey. In bacteria, an ecologically distinct population (called an “ecotype”) may constitute a separate species, but only if the distinction is permanent. As evolutionary microbiologist Frederick Cohan wrote in 2002, species in bacteria “are ecologically distinct from one another; and they are irreversibly separate.”47 The rapid reversal of ecological distinctions when the bacterial populations in Rainey’s experiment were put back into the same environment refutes Coyne’s claim that the experiment demonstrated the origin of a new species.
Exaggerating the evidence to prop up Darwinism is not new. In the Galápagos finches, average beak depth reverted to normal after the drought ended. There was no net evolution, much less speciation. Yet Coyne writes in Why Evolution Is True that “everything we require of evolution by natural selection was amply documented” by the finch studies. Since scientific theories stand or fall on the evidence, Coyne’s tendency to exaggerate the evidence does not speak well for the theory he is defending. When a 1999 booklet published by The U. S. National Academy of Sciences called the change in finch beaks “a particularly compelling example of speciation,” Berkeley law professor and Darwin critic Phillip E. Johnson wrote in The Wall Street Journal: “When our leading scientists have to resort to the sort of distortion that would land a stock promoter in jail, you know they are in trouble.”48
So there are observed instances of secondary speciation — which is not what Darwinism needs — but no observed instances of primary speciation, not even in bacteria. British bacteriologist Alan H. Linton looked for confirmed reports of primary speciation and concluded in 2001: “None exists in the literature claiming that one species has been shown to evolve into another. Bacteria, the simplest form of independent life, are ideal for this kind of study, with generation times of twenty to thirty minutes, and populations achieved after eighteen hours. But throughout 150 years of the science of bacteriology, there is no evidence that one species of bacteria has changed into another.”49
Conclusions
Darwin called The Origin of Species “one long argument” for his theory, but Jerry Coyne has given us one long bluff. Why Evolution Is True tries to defend Darwinian evolution by rearranging the fossil record; by misrepresenting the development of vertebrate embryos; by ignoring evidence for the functionality of allegedly vestigial organs and non-coding DNA, then propping up Darwinism with theological arguments about “bad design;” by attributing some biogeographical patterns to convergence due to the supposedly “well-known” processes of natural selection and speciation; and then exaggerating the evidence for selection and speciation to make it seem as though they could accomplish what Darwinism requires of them.
The actual evidence shows that major features of the fossil record are an embarrassment to Darwinian evolution; that early development in vertebrate embryos is more consistent with separate origins than with common ancestry; that non-coding DNA is fully functional, contrary to neo-Darwinian predictions; and that natural selection can accomplish nothing more than artificial selection — which is to say, minor changes within existing species.
Faced with such evidence, any other scientific theory would probably have been abandoned long ago. Judged by the normal criteria of empirical science, Darwinism is false. Its persists in spite of the evidence, and the eagerness of Darwin and his followers to defend it with theological arguments about creation and design suggests that its persistence has nothing to do with science at all.50
Nevertheless, biology students might find Coyne’s book useful. Given accurate information and the freedom to exercise critical thinking, students could learn from Why Evolution Is True how Darwinists manipulate the evidence and mix it with theology to recycle a false theory that should have been discarded long ago.
Notes
- Jerry A. Coyne, Why Evolution Is True (New York: Viking, 2009), p. 3.
- Coyne, Why Evolution Is True, pp. 3-4.
- Coyne, Why Evolution Is True, pp. 5-6.
- Coyne, Why Evolution Is True, pp. 18-19.
- Coyne, Why Evolution Is True, pp. 17-18, 25.
- Charles Darwin, The Origin of Species, Sixth Edition (London: John Murray, 1872), Chapter X, pp. 266, 285-288. Available online (2009) here.
- J. William Schopf, “The early evolution of life: solution to Darwin’s dilemma,” Trends in Ecology and Evolution 9 (1994): 375-377.
James W. Valentine, Stanley M. Awramik, Philip W. Signor & M. Sadler, “The Biological Explosion at the Precambrian-Cambrian Boundary,” Evolutionary Biology 25 (1991): 279-356.
James W. Valentine & Douglas H. Erwin, “Interpreting Great Developmental Experiments: The Fossil Record,” pp. 71-107 in Rudolf A. Raff & Elizabeth C. Raff, (editors), Development as an Evolutionary Process (New York: Alan R. Liss, 1987).
Jeffrey S. Levinton, “The Big Bang of Animal Evolution,” Scientific American 267 (November, 1992): 84-91.
“The Scientific Controversy Over the Cambrian Explosion,” Discovery Institute. Available online (2009) here.
Jonathan Wells, Icons of Evolution (Washington, DC: Regnery Publishing, 2002), Chapter 3. More information available online (2009) here.
Stephen C. Meyer, “The Cambrian Explosion: Biology’s Big Bang,” pp. 323-402 in John Angus Campbell & Stephen C. Meyer (editors), Darwinism, Design, and Public Education (East Lansing, MI: Michigan State University Press, 2003). More information available online (2009) here. - Coyne, Why Evolution Is True, p. 28.
- Coyne, Why Evolution Is True, p. 48.
- Coyne, Why Evolution Is True, pp. 49-51.
- Kevin Padian & Luis M. Chiappe, “The origin and early evolution of birds,” Biological Reviews 73 (1998): 1-42. Available online (2009) here.
Wells, Icons of Evolution, pp. 119-122. - Coyne, Why Evolution Is True, pp. 25, 53.
Chico Marx in Duck Soup (Paramount Pictures, 1933). This and other Marx Brothers quotations are available online (2009) here. - Gareth Nelson, “Presentation to the American Museum of Natural History (1969),” in David M. Williams & Malte C. Ebach, “The reform of palaeontology and the rise of biogeography—25 years after ‘ontogeny, phylogeny, palaeontology and the biogenetic law’ (Nelson, 1978),” Journal of Biogeography 31 (2004): 685-712.
- Henry Gee, In Search of Deep Time. New York: Free Press, 1999, pp. 5, 32, 113-117.
Jonathan Wells, The Politically Incorrect Guide to Darwinism and Intelligent Design (Washington, DC: Regnery Publishing, 2006). More information available online (2009) here. - Coyne, Why Evolution Is True, p. 79.
Darwin, The Origin of Species, Chapter XIV, pp. 386-396. Available online (2009) here. - Darwin, The Origin of Species, Chapter XIV, pp. 387-388. Available online (2009) here.
- Coyne, Why Evolution Is True, p. 73.
Karl Ernst von Baer, “On the Development of Animals, with Observations and Reflections: The Fifth Scholium,” translated by Thomas Henry Huxley, pp. 186-237 in Arthur Henfrey & Thomas H. Huxley (editors), Scientific Memoirs: Selected from the Transactions of Foreign Academies of Science and from Foreign Journals: Natural History (London, 1853; reprinted 1966 by Johnson Reprint Corporation, New York). The passage quoted by Darwin is on p. 210.
Jane M. Oppenheimer, “An Embryological Enigma in the Origin of Species,” pp. 221-255 in Jane M. Oppenheimer, Essays in the History of Embryology and Biology (Cambridge, MA: The M.I.T. Press, 1967). - Timothy Lenoir, The Strategy of Life (Chicago: The University of Chicago Press, 1982), p. 258.
Frederick B. Churchill, “The Rise of Classical Descriptive Embryology,” pp. 1-29 in Scott F. Gilbert (editor), A Conceptual History of Modern Embryology (Baltimore, MD: The Johns Hopkins University Press, 1991), pp. 19-20. - Coyne, Why Evolution Is True, pp. 77-79.
- Simon Conway Morris, “Fossil Embryos,” pp. 703-711 in Claudio D. Stern (editor), Gastrulation: From Cells to Embryos (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 2004).
- Walter Garstang, “The theory of recapitulation: a critical restatement of the biogenetic law,” Journal of the Linnean Society (Zoology), 35 (1922): 81-101.
- See Chapter Five and accompanying references in Wells, Icons of Evolution.
See Chapter Three and accompanying references in Wells, The Politically Incorrect Guide to Darwinism and Intelligent Design. - Michael K. Richardson, J. Hanken, M. L. Gooneratne, C. Pieau, A. Raynaud, L. Selwood & G. M. Wright, “There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development,” Anatomy & Embryology 196 (1997): 91-106.
Michael K. Richardson, quoted in Elizabeth Pennisi, “Haeckel’s Embryos: Fraud Rediscovered,” Science 277 (1997): 1435.
Stephen Jay Gould, “Abscheulich! Atrocious!” Natural History (March, 2000), pp. 42-49.
“Hoax of Dodos” (2007). Available online (2009) here. - Coyne, Why Evolution Is True, p. 78.Notes
- Darwin, The Origin of Species, Chapters XIV (p. 402) and XV (p. 420). Available online (2009) here.
- Darwin, Charles. The Descent of Man, First Edition (London: John Murray, 1871), Chapter I (p. 27). Available online (2009) here.
Kohtaro Fujihashi, J.R. McGhee, C. Lue, K.W. Beagley, T. Taga, T. Hirano, T. Kishimoto, J. Mestecky & H. Kiyono, “Human Appendix B Cells Naturally Express Receptors for and Respond to Interleukin 6 with Selective IgA1 and IgA2 Synthesis,” Journal of Clinical Investigations 88 (1991): 248-252. Available online (2009) here.
J.A. Laissue, B.B. Chappuis, C. Müller, J.C. Reubi & J.O. Gebbers, “The intestinal immune system and its relation to disease,” Digestive Diseases (Basel) 11 (1993): 298-312. Abstract available online (2009) here.
Loren G. Martin, “What is the function of the human appendix?” Scientific American (October 21, 1999), Available online (2009) here.
R. Randal Bollinger, Andrew S. Barbas, Errol L. Bush, Shu S. Lin & William Parker, “Biofilms in the large bowel suggest an apparent function of the human vermiform appendix,” Journal of Theoretical Biology 249 (2007): 826-831. Available online (2009) here.
Duke University Medical Center, “Appendix Isn’t Useless At All: It’s A Safe House For Good Bacteria,” ScienceDaily (October 8, 2007). Available online (2009) here. - Steven R. Scadding, “Do ‘vestigial organs’ provide evidence for evolution?” Evolutionary Theory 5 (1981): 173-176.
Bruce G. Naylor, “Vestigial organs are evidence of evolution,” Evolutionary Theory 6 (1982): 91-96.
Steven R. Scadding, “Vestigial organs do not provide scientific evidence for evolution,” Evolutionary Theory 6 (1982): 171-173. - Coyne, Why Evolution Is True, pp. 61-62.
- Coyne, Why Evolution Is True, p. 46.
- Coyne, Why Evolution Is True, pp. 81.
- Susumu Ohno, “So much ‘junk’ DNA in our genome,” Brookhaven Symposia in Biology 23 (1972): 366-70.
Richard Dawkins, The Selfish Gene (New York: Oxford University Press, 1976), p. 47. - Coyne, Why Evolution Is True, pp. 66-67.
- A few of the many scientific articles published since 2003 that document the function of so-called “junk” DNA are:
- E.S Balakirev & F.J. Ayala, “Pseudogenes: are they ‘junk’ or functional DNA?” Annual Review of Genetics 37 (2003): 123-151.
- A. Hüttenhofer, P. Schattner & N. Polacek, “Non-coding RNAs: hope or hype?” Trends in Genetics 21 (2005): 289-297.
- J.S. Mattick & I.V. Makunin, “Non-coding RNA,” Human Molecular Genetics 15 (2006): R17-R29.
- R.K. Slotkin & R. Martienssen, “Transposable elements and the epigenetic regulation of the genome,” Nature Reviews Genetics 8 (2007): 272-285.
- P. Carninci, J. Yasuda & Y Hayashizaki, “Multifaceted mammalian transcriptome,” Current Opinion in Cell Biology 20 (2008): 274-80.
- C.D. Malone & G.J. Hannon, “Small RNAs as Guardians of the Genome,” Cell 136 (2009): 656–668.
- C.P. Ponting, P.L. Oliver & W. Reik, “Evolution and Functions of Long Noncoding RNAs,” Cell 136 (2009): 629–641.
- Darwin, The Origin of Species, Chapters XIII (pp. 347-352) and XV (p. 419). Available online (2009) here.
- Darwin, The Origin of Species, Chapters XII (pp. 330-332). Available online (2009) here.
- Alan Cooper, et al., C. Mourer-Chauviré, C.K. Chambers, A. von Haeseler, A.C. Wilson & S. Paabo, “Independent origins of New Zealand moas and kiwis,” Proceedings of the National Academy of Sciences USA 89 (1992): 8741-8744. Available online (2008) here.
Oliver Haddrath & Allan J. Baker, “Complete mitochondrial DNA genome sequences of extinct birds: ratite phylogenetics and the vicariance biogeography hypothesis,” Proceedings of the Royal Society of London B 268 (2001): 939-945.
John Harshman, E.L. Braun, M.J. Braun, C.J. Huddleston, R.C.K. Bowie,
J.L. Chojnowski, S.J. Hackett, K.-L. Han, R.T. Kimball, B.D. Marks, K.J. Miglia,
W.S. Moore, S. Reddy, F.H. Sheldon, D.W. Steadman, S.J. Steppan, C.C. Witt & T. Yuri, “Phylogenomic evidence for multiple losses of flight in ratite birds,” Proceedings of the National Academy of Sciences USA 105 (2008): 13462-13467. Abstract available online (2008) here.
Giuseppe Sermonti, “L’evoluzione in Italia – La via torinese / How Evolution Came to Italy – The Turin Connection,” Rivista di Biologia/Biology Forum 94 (2001): 5-12. Available online (2008) here. - Giuseppe Colosi, “La distribuzione geografica dei Potamonidae,” Rivista di Biologia 3 (1921): 294-301. Available online (2009) here.
Savel R. Daniels, N. Cumberlidge, M. Pérez-Losada, S.A.E. Marijnissen &
K.A. Crandall, “Evolution of Afrotropical freshwater crab lineages obscured by morphological convergence,” Molecular Phylogenetics and Evolution 40 (2006): 227–235. Available online (2009) here.
R. von Sternberg, N. Cumberlidge & G. Rodriguez. “On the marine sister groups of the freshwater crabs (Crustacea: Decapoda: Brachyura),” Journal of Zoological Systematics and Evolutionary Research 37 (1999): 19–38.
Darren C.J. Yeo, et al., “Global diversity of crabs (Crustacea: Decapoda: Brachyura) in freshwater,” Hydrobiologia 595 (2008): 275-286. - Léon Croizat, Space, Time, Form: The Biological Synthesis. Published by the author (Deventer, Netherlands: N. V. Drukkerij Salland, 1962), p. iii.
Robin C. Craw, “Léon Croizat’s Biogeographic Work: A Personal Appreciation,” Tuatara 27:1 (August 1984): 8-13. Available online (2009) here.
John R. Grehan, “Evolution By Law: Croizat’s ‘Orthogeny’ and Darwin’s ‘Laws of Growth’,” Tuatara 27:1 (August 1984): 14-19. Available online (2009) here.
Carmen Colacino, “Léon Croizat’s Biogeography and Macroevolution, or… ‘Out of Nothing, Nothing Comes’,” The Philippine Scientist 34 (1997): 73-88.
Ernst Mayr, The Growth of Biological Thought (Cambridge, MA: Harvard University Press, 1982), pp. 529-530. - Coyne, Why Evolution Is True, pp. 92-94.
- Coyne, Why Evolution Is True, p. 116.
Darwin, The Origin of Species, Chapter IV (p. 70). Available online (2009) here.
H. B. D. Kettlewell, “Darwin’s Missing Evidence,” Scientific American 200 (March, 1959): 48-53. - Ernst Mayr, The Growth of Biological Thought (Cambridge, MA: Harvard University Press, 1982), p. 403.
Ernst Mayr, Populations, Species and Evolution (Cambridge, MA: Harvard University Press, 1963), p. 10.
Keith Stewart Thomson, “Natural Selection and Evolution’s Smoking Gun,” American Scientist 85 (1997): 516-518. - Jerry A. Coyne & H. Allen Orr, Speciation (Sunderland, MA: Sinauer Associates, 2004), p. 25-39.
Coyne, Why Evolution Is True, p. 174. - Coyne, Why Evolution Is True, p. 188.
- Douglas J. Futuyma, Evolution (Sunderland, MA: Sinauer Associates, 2005), p. 398.
- Wells, The Politically Incorrect Guide to Darwinism and Intelligent Design, Chapter Five (“The Ultimate Missing Link”), pp. 49-59.
- Coyne, Why Evolution Is True, pp. 129-130.
- Paul B. Rainey & Michael Travisano. “Adaptive radiation in a heterogeneous environment,” Nature 394 (1998): 69-72.
Frederick M. Cohan, “What Are Bacterial Species?” Annual Review of Microbiology 56 (2002): 457-482. Available online (2009) here. - Coyne, Why Evolution Is True, p. 134.
National Academy of Sciences, Science and Creationism: A View from the National Academy of Sciences, Second edition (Washington, DC: National Academy of Sciences Press, 1999), Chapter on “Evidence Supporting Biological Evolution,” p. 10. Available online (2009) here.
Phillip E. Johnson, “The Church of Darwin,” The Wall Street Journal (August 16, 1999): A14. Available online (2009) here. - Alan H. Linton, “Scant Search for the Maker,” The Times Higher Education Supplement (April 20, 2001), Book Section, p. 29.
Frederick M. Cohan, “What Are Bacterial Species?” Annual Review of Microbiology 56 (2002): 457-482. Available online (2009) here. - Paul A. Nelson, “The role of theology in current evolutionary reasoning,” Biology and Philosophy 11 (October 1996): 493 – 517. Abstract available online (2009) here.
Jonathan Wells, “Darwin’s Straw God Argument,” Discovery Institute (December 2008). Available online (2009) here. Jonathan Wells, “Darwin’s Straw God Argument,” Discovery Institute (December 2008). Available online (2009) here.