
A Mousetrap Defended
Response to CriticsIntroduction
In Darwin’s Black Box: The Biochemical Challenge to Evolution I coined the term “irreducible complexity” in order to point out an apparent problem for the Darwinian evolution of some biochemical and cellular systems. In brief, an irreducibly complex system is one that needs several well-matched parts, all working together, to perform its function. The reason that such systems are headaches for Darwinism is that it is a gradualistic theory, wherein improvements can only be made step by tiny step,1 with no thought for their future utility. I argued that a number of biochemical systems, such as the blood clotting cascade, intracellular transport system, and bacterial flagellum are irreducibly complex and therefore recalcitrant to gradual construction, and so they fit poorly within a Darwinian framework. Instead I argued they are best explained as the products of deliberate intelligent design.
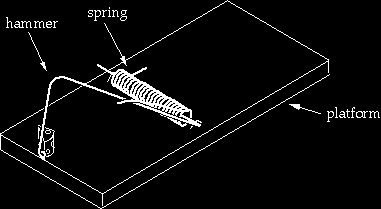
In order to communicate the concept to a general audience, I used a mousetrap as an example of an irreducibly complex system in everyday life. The mousetrap I pictured in my book had a number of parts that all had to work together to catch mice. The usefulness of the mousetrap example was that it captured the essence of the problem I saw for gradualistic evolution at a level that could be understood by people who were unfamiliar with the fine points of protein structure and function — that is, nearly everyone. For that same reason, defenders of Darwinism have assailed it. Although it may seem silly to argue over a mousetrap, it is actually critical to allowing people who are not professional scientists to understand the issues involved. In this article I defend the mousetrap as an example of irreducible complexity that can’t be put together by a series of small, undirected steps.
Mousetrap rebuttals have popped up in a variety of situations including national television, but most recently (June 2000) was at a conference I attended at Concordia University in Wisconsin where Kenneth Miller, professor of biology at Brown University, spent several minutes during his presentation attacking the mousetrap. In doing so he used images of mousetraps that were drawn by Professor John McDonald of the University of Delaware and can be seen on his web site2 (reproduced below with permission). In defense of the mousetrap I will make a number of points, including: (1) McDonald’s reduced-component traps are not single-step intermediates in the building of the mousetrap I showed; (2) intelligence was intimately involved in constructing the series of traps; 3 if intelligence is necessary to make something as simple as a mousetrap, we have strong reason to think it is necessary to make the much more complicated machinery of the cell.
Conceptual precursors vs. physical precursors
On his web site Professor McDonald was careful to make a critical distinction. He clearly stated “the reduced-complexity mousetraps . . . are intended to point out the logical flaw in the intelligent design argument; they’re not intended as an analogy of how evolution works.” Nonetheless Kenneth Miller discussed McDonald’s examples in a way that would lead an audience to think that they were indeed relevant to Darwinian evolution. Only at the end of the presentation did he briefly mention the disanalogy. I believe such tactics are disingenuous at best, like tagging a brief warning onto the end of a cigarette commercial containing attractive images. The purpose of the images is to get you to buy the cigarettes, despite the warning. The purpose of citing McDonald’s drawings is to get people to buy Darwinian evolution, despite the brief disclaimer.
The logical point Professor McDonald wished to make was that there are mousetraps that can work with fewer parts than the trap I pictured in my book. Let me say that I agree completely; in fact, I said so in my book (see below). For example, one can dig a steep hole in the ground for mice to fall into and starve to death. Arguably that has zero parts. One can catch mice with a glue trap, which has only one part. One can prop up a box with a stick, hoping a mouse will bump the stick and the box will fall on top of it. That has two parts. And so forth. There is no end to possible variation in mousetrap design. But, as I tried to emphasize in my book, the point that is relevant to Darwinian evolution is not whether one can make variant structures, but whether those structures lead, step-by-excruciatingly-tedious-Darwinian-step, to the structure I showed. I wrote(3):
To feel the full force of the conclusion that a system is irreducibly complex and therefore has no functional precursors we need to distinguish between a physical precursor and a conceptual precursor. The trap described above is not the only system that can immobilize a mouse. On other occasions my family has used a glue trap. In theory at least, one can use a box propped open with a stick that could be tripped. Or one can simply shoot the mouse with a BB gun. However, these are not physical precursors to the standard mousetrap since they cannot be transformed, step-by-Darwinian-step, into a trap with a base, hammer, spring, catch, and holding bar.
Since I agree with Professor McDonald that there could be mousetraps with fewer parts, the only relevant question is whether the mousetraps he drew are physical precursors, or merely conceptual precursors. Can they “be transformed, step-by-Darwinian-step” into the trap I pictured (essentially the same structure as the fifth trap shown below), as some people have been led to believe? No, they can’t.
From the first trap to the second
Professor McDonald started with a complete mousetrap and then showed ones with fewer parts. I will reverse that order, start with his simplest trap, and show the steps that would be necessary to convert it into the next more complex trap in his series. That, after all, is the way Darwinian evolution would have to work. If we are to picture this as a Darwinian process, then each separate adjustment must count as a “mutation.” If several separate mutations have to occur before we go from one functional trap to the next, then a Darwinian process is effectively ruled out, because the probability of getting multiple unselected mutations that eventually lead to a specific complex structure is prohibitive. Shown below are the simplest and next-to-simplest traps.
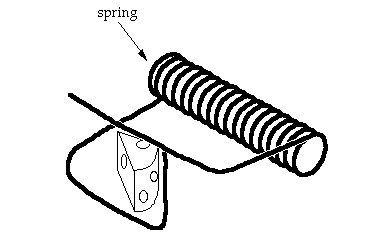
The single-piece trap, consisting of just a spring with extended arms, is supposed to have one arm, under tension, propped up on the other arm. When a mouse jiggles it, the arm is released and comes down, pinning the mouse’s paw against the other arm. Now, the first thing to notice is that the single piece trap isn’t a simple spring — it’s got a very specific structure. If the lengths of the extended ends varied by much before their first bend, or if the angle of the bends differed somewhat, the trap wouldn’t work. What’s more, the strength of the material out of which the spring is made has to be consonant with the purpose of catching a mouse (for example, if it were made from an old Slinky it likely wouldn’t work). It is not a simple starting point; it was intelligently selected. Nonetheless, I realize that in coming up with an analogy we have to start somewhere. So I will not complain about an intelligently-selected starting point. However, the involvement of intelligence at any other point along the way invalidates the entire exercise as an analogy to a Darwinian process. Because Darwinism wholly rejects intelligent direction, Darwinists must agree that the involvement of intelligence at any point in a scenario (after the agreed-on starting point) is fatal. That point occurs immediately for our mousetrap.
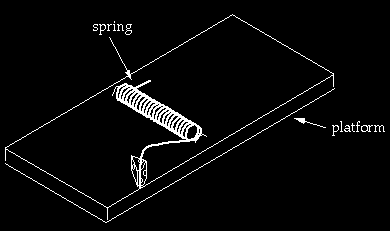
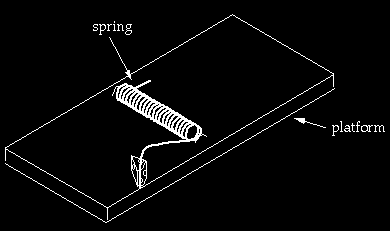
The second mousetrap (above) has a spring and a platform. One of the extended arms stands under tension at the very edge of the platform. The idea is that if a mouse in the vicinity jiggles the trap, the end of the arm slips over the edge and comes rushing down, and may pin the mouse’s paw or tail against the platform. Now, the first thing to notice is that the arms of the spring are in a different relationship to each other than in the first trap. To get to the configuration of the spring in the second trap from the configuration in the first, it seems to me one would have to proceed through the following steps (4): (1) twist the arm that has one bend through about 90* so that the end segment is perpendicular to the axis of the spring and points toward the platform; (2) twist the other arm through about 180* so the first segment is pointing opposite to where it originally pointed (the exact value of the rotations depend on the lengths of the arms); (3) shorten one arm so that its length is less than the distance from the top of the platform to the floor (so that the end doesn’t first hit the floor before pinning the mouse). While the arms were being rotated and adjusted, the original one-piece trap would have lost function, and the second trap would not yet be working.
At this point we bring in a new piece, the platform, which is a simple piece of wood. One now has a spring resting on top of a platform. However, the spring cannot be under tension in this configuration unless it is fixed in place. Notice that in the second mousetrap, not only has a platform been added, but two (barely visible) staples have been added as well. Thus we have gone not from a one piece to a two-piece trap, but from a one to a four piece trap. Two staples are needed; if there were only one staple positioned as drawn, the tensed spring would be able to rotate out of position. The staples have to be positioned carefully with respect to the platform. They have to be arranged within a very narrow tolerance so that one arm of the spring teeters perilously on the edge of the platform or the trap doesn’t work. If either of the staples is moved significantly from where they are drawn, the trap won’t function. I should add that I did not emphasize the staples in my book because I was trying to make a simple point and didn’t want to exhaust the readers with tedium. However, someone who wishes to seriously propose that the mousetrap I pictured is approachable in the tiny steps required by Darwinian processes would indeed have to deal with all the details, including the staples.
It is important to remember that the placement, size, shape, or any important feature (not just “piece”) of a system can’t just be chosen to fit the purposes of a person who wishes to simulate a Darwinian process. Rather, each significant feature has to be justified as being a small improvement. In the real world the occasional unselected feature might occur which serendipitously will be useful in the future, but invoking more than one unselected (neutral, nonadaptive) event in a Darwinian scenario seems to me impermissible because the improbability of the joint events starts to soar. In our current case the unselected event we are allowed was used up when we began with a special starting point.
I think the problems of rearranging the already-functioning first mousetrap shows the general difficulties one expects in trying to re-arrange an already-functioning system into something else. The requirements (“selection pressures”) that make a component suitable for one specialized system will generally make it unsuitable for another system without significant modification. Another problem we can note is that the second mousetrap is not an obvious improvement over the first; it is difficult to see how it would function any better than the one-piece trap. It’s just that it’s on the road to where we want to see the system end up — on the road to a distant target. That, of course, is intelligent direction.
The transition from the first to the second mousetrap is not analogous to a Darwinian process because: (1) a number of separate steps are required to make the transition; (2) each step has to fall within a narrow range of tolerance to get to the target trap; and (3) function is lost until the transition is completed. In fact, the situation of going from the first trap to the second trap is best viewed not as a transition, but as building a different kind of trap using some old materials from the first trap (with major modifications) and some new materials. Far from being an analogy to a Darwinian process, the construction of the second trap is an example of intelligent design.
From the second trap to the third
The way the traps are drawn (below), the transition from the second to the third trap doesn’t seem to be a big step. Both drawings are superficially similar. But when one thinks about the transition in detail problems crop up. The first problem is that a new piece is added — the hammer. Unlike the platform that was added in the last transition (which I did not object to), the hammer is not a simple object. Rather it contains several bends. The angles of the bends have to be within relatively narrow tolerances for the end of the hammer to be positioned precisely at the edge of the platform, otherwise the system doesn’t work. For the same reason, the length of the second segment of the hammer has to be within a narrow range of values. How does the hammer get into the third trap? It would seem that the extended arm of the second trap has to be held up while the newly-fashioned hammer is inserted through the tunnel of the spring. Thus an intelligent agent has to actively push parts around to get to the configuration of the third trap. Again, there is no obvious improvement in function of the third trap compared to the second or first. Both the second and third traps appear to do the same thing as the first, but require more parts. Such an event is not expected in a Darwinian scenario. It seems the only reason they attract our attention is because they appear to be along the path we wish the process would go. That is intelligent design.
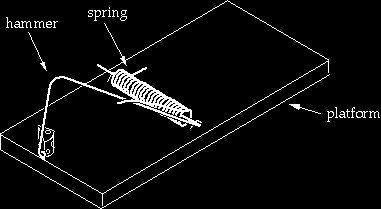
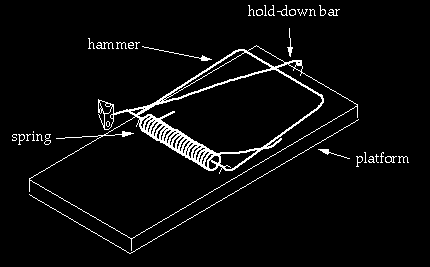
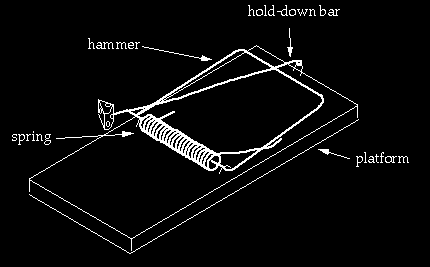
From the third trap to the fourth
Going from the third trap to the fourth requires major rearrangements. The hammer is bent, lengthened, and an extra segment is added to it. Two new pieces are added: the “hold-down bar” and a staple to hold down the hold-down bar. The end of the hold-down bar is endowed with a closed curl so that the staple has something to hang on to. The staple again has to be positioned in a specific region of the trap. Depending on details, this configuration may be an improvement over the first three traps because it appears that, depending on the tension of the spring, the trap could kill a mouse outright, rather than just pinning it (yet that feature could probably easily be built into the earlier versions). On the other hand the arm of the spring is now being pushed through a much greater displacement in the fourth trap than previous versions. It seems unlikely a spring optimized for use in earlier traps would work well in the fourth trap (unless of course we are “looking ahead”). Rather than a “transition,” this process is again better viewed as building a new trap using refashioned parts of the old trap plus new ones. This is intelligent design.
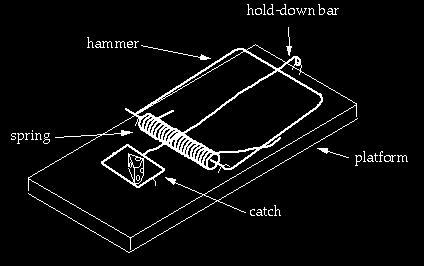
From the fourth trap to the fifth
This is left as an exercise for the reader.
Discussion
I have to admit that even I find it tedious to discuss mousetraps in such excruciating detail. But the critical point is that that is exactly the level at which Darwinian evolution would have to work in the cell. Every relevant detail has to fit or the system fails. If an arm is too long or an angle not right or a staple placed incorrectly, the mouse dances free. If you want to get to a certain system, but the road there isn’t a series of continual improvements, Darwinism won’t take you there. It’s important for those interested in these issues to realize that, when evaluating descriptive evolutionary scenarios (as opposed to experiments — see below), one has to attend to the tiniest details (as I did here) to see if intelligence is directing the show. On the other hand, if one doesn’t pay the strictest attention, Darwinian scenarios look much more plausible because one sees only the possibilities, not the problems. It’s easy for a speaker to persuade an audience that the McDonald mousetraps represent a series of Darwinian intermediates on the way to a standard trap — that they show irreducible complexity is no big deal. All one has to do is gloss over the difficulties. But although our minds can skip over details, nature can’t.
In the real world of biology the staples, bends, and so forth would be features of molecules, of proteins in particular. If two proteins don’t bind each other in the correct orientation (aren’t stapled right), if they aren’t placed in the right positions, if their new activity isn’t regulated correctly, if many details aren’t exactly correct, then the putative Darwinian pathway is blocked. Now, it’s hard, almost impossible, for persons without the appropriate science background to tell where such difficulties would occur in Darwinian scenarios for blood clotting or ciliary function or other biological systems. When they read Darwinian stories in a book or hear them in lectures, they generally have no independent information to judge the scenario. In such a situation one should ask oneself, “If a simple mousetrap requires intelligent design, what is the likelihood that the much more complicated molecular machines of the cell could be built step-by-tiny-Darwinian-step?” Keeping that question in mind will foster a healthy skepticism toward optimistic scenarios.
Why do the McDonald mousetraps look persuasive to some people? Certainly one reason is the way they are drawn. Drawings of four of the five traps are dominated by the image of the large rectangular platform and prominent spring in the center. That makes them all look pretty much the same. The staples are barely visible and the various metal bars protruding here and there seem like insignificant details. In fact, they are critical. Another reason is that the scenario starts with the completed mousetrap. Any question about the placement of the parts, their size, stiffness, and so on doesn’t easily arise because the parts were already placed where they needed to be for the ultimate goal in the original drawing (that is, the fifth mousetrap here, which is the first drawing in McDonald’s series) and their properties could be inferred from the fact we started with a working trap. The universe of possibilities was tightly but implicitly circumscribed by the already-completed starting point. A third reason it seems persuasive is that the series is always presented as parts being removed from the complete mousetrap. Looking at it in such a backward manner — the reverse of what evolution would have to do — obscures the teleology of the building process. Going in a forward direction there is strong reason to think we would not end up at the fifth mousetrap when starting from the first, because the first works as well as the second and third, so greater complexity would be disfavored. In going backwards, however, lesser complexity is favored so it seems “natural” to move to simpler traps. Yet Darwinian evolution can’t work like that.
A final reason for the persuasiveness of the example we can call the “Clever Hans effect.” Clever Hans was the name of a horse who seemed to be pretty good at arithmetic. Its owner would give Hans a simple math problem such as 5+5, and the horse would stamp his hoof ten times, then stop. It eventually turned out that Clever Hans could pick up unconscious cues from its owner, who might raise his eyebrows or tilt his head when the horse’s stamping reached the right value. The horse could even pick up unintentional cues from other people, not just the owner, who also apparently gave telltale reactions. In the case of Clever Hans, the human intelligence of the owner was inadvertently attributed to the horse. In my experience the same is invariably true of Darwinian scenarios — human intelligence is critical to guiding the scenario through difficulties toward the “proper” goal, but the intelligence is then attributed to natural selection. As with Clever Hans, the guidance is usually unconscious, but is intelligent nonetheless.
Clever Hans was exposed as mathematically clueless by carefully controlled experiments. To see whether natural selection can work wonders on its own — without the aid of human intelligence — we also have to do carefully controlled experiments. One way to do this is to ask bacteria in the laboratory if they can evolve irreducibly complex biochemical systems. (Kenneth Miller has called this the “acid test.”) Bacteria are a good choice because they can be grown in huge numbers with short generation times — just what Darwinian evolution needs. However, when this was repeatedly tried over the course of 25 years for bacteria missing a comparatively simple biochemical system (called the “lac operon”) natural selection came up empty (see “The Acid Test” on this website). It could make the small changes typically termed “microevolution,” but whenever it had to do a couple things at once, such as would have to be done to make irreducibly complex systems, it got stuck.(5) Like Clever Hans on his own, natural selection seems to have much less intelligence than we had given it credit for. There is currently no experimental evidence to show that natural selection can get around irreducible complexity.
Darwinian scenarios, either for building mousetraps or biochemical systems, are very easy to believe if we aren’t willing or able to scrutinize the smallest details, or to ask for experimental evidence. They invite us to admire the intelligence of natural selection. But the intelligence we are admiring is our own.
Endnotes
- One has to be sophisticated about what is regarded as a “step.” One mutational step in a biological organism might seem to have large effects, such as the famous antennapedia mutation in fruit flies. Although such a change may impress us, it involves only the rearrangement of existing structures; no new structures are made. When thinking about what’s involved in making a new structure it’s best to think of how many lines of instructions (analogous to lines of computer code) would be needed to build it. See my discussion of this topic in Darwin’s Black Box, pp. 39-41.
- http://udel.edu/~mcdonald/mousetrap.html
- Behe, M. J. (1996). Darwin’s black box: the biochemical challenge to evolution. (The Free Press: New York), p. 43.
- To play the game right, one has to compare the probability of these events happening with the probability of any slight “mutation” happening. To give a flavor of what that might mean, a mutation might involve bending the spring in the middle, changing the size of the platform, changing the tension on the spring, extending the end of a metal piece, and so on. A crude feel for the probabilities of the events can be obtained by examining the precision a feature must have for the trap to work. To get the probability for two or more unselected events (ones that don’t improve the function), one multiplies the probabilities for each.
- The work has been done by Barry Hall of the University of Rochester. He could get selection to replace one piece of the lac operon (the B-galactosidase) but had to intelligently intervene to keep the bacteria alive by adding the artificial chemical induce IPTG. The bacteria could not replace two required protein parts deleted simultaneously, showing the severe problem of irreducible complexity. For a review of his work see: Hall, B. G. (1999). Experimental evolution of Ebg enzyme provides clues about the evolution of catalysis and to evolutionary potential. FEMS Microbiology Letters 174, 1-8.